

ADC在癌症治疗中的分子与免疫学驱动机制
ADC在癌症治疗中的分子与免疫学驱动机制的核心信息是什么?
研究背景 抗体偶联药物(ADC)通过将抗体的靶向精准性与细胞毒性药物的强效杀伤相结合,已成为癌症治疗的重要方向。近期, Nature Reviews Cancer 杂志发布综述,系统阐述了ADC的作用机制,为深入理解其作用原理与优化临床策略提供了关键见解。【ONCO前沿】特此编译,以飨读者。 研究封面 ADC的经典结构由单克隆抗体(主要为IgG型)通过连接子...
研究背景 抗体偶联药物(ADC)通过将抗体的靶向精准性与细胞毒性药物的强效杀伤相结合,已成为癌症治疗的重要方向。近期, Nature Reviews Cancer 杂志发布综述,系统阐述了ADC的作用机制,为深入理解其作用原理与优化临床策略提供了关键见解。【ONCO前沿】特此编译,以飨读者。 属于「ONCO前沿」分类。 关键词:抗体偶联药物、抗肿瘤药物、免疫肿瘤学。
本文核心问答
Q1: 这篇资讯的核心内容是什么?
研究背景 抗体偶联药物(ADC)通过将抗体的靶向精准性与细胞毒性药物的强效杀伤相结合,已成为癌症治疗的重要方向。近期, Nature Reviews Cancer 杂志发布综述,系统阐述了ADC的作用机制,为深入理解其作用原理与优化临床策略提供了关键见解。【ONCO前沿】特此编译,以飨读者。
Q2: 本文属于哪个学科分类,涉及哪些关键词?
本文属于「ONCO前沿」分类,涉及关键词:抗体偶联药物、抗肿瘤药物、免疫肿瘤学。
Q3: 这篇资讯的发布时间是什么?
本文发布于1766655066,来源为TalkMED拓麦医学资讯平台。

研究背景
抗体偶联药物(ADC)通过将抗体的靶向精准性与细胞毒性药物的强效杀伤相结合,已成为癌症治疗的重要方向。近期,Nature Reviews Cancer杂志发布综述,系统阐述了ADC的作用机制,为深入理解其作用原理与优化临床策略提供了关键见解。【ONCO前沿】特此编译,以飨读者。

研究封面
ADC的经典结构由单克隆抗体(主要为IgG型)通过连接子与小分子药物(通常为细胞毒性有效载荷)相连而成。这些组分并非独立发挥作用,而是通过与全身各组织细胞(包括肿瘤细胞)的复杂相互作用,共同决定ADC的疗效与毒性特征。
鉴于ADC设计的复杂性,该综述将分为三部分进行阐述。第一部分聚焦ADC在体内的分布及其与疗效的关系。从血液中的系统浓度开始,依次介绍其在肿瘤中的摄取、在肿瘤微环境(TME)中的分布,以及导致有效载荷释放的细胞和亚细胞处理过程。细胞毒性有效载荷的递送是驱动这类药物疗效的主要(但非唯一)作用机制。第二部分重点探讨ADC的毒性。由于全身递送的固有局限性,多数ADC并未定位于肿瘤。其在非肿瘤组织中的代谢与有效载荷释放,以及这些组织对有效载荷的敏感性,很大程度上构成了ADC的特异性毒性谱。该部分还将分析影响分布的关键因素,如连接子稳定性和抗体设计,这些因素对减少脱靶毒性至关重要。第三部分将讨论ADC与免疫系统的相互作用。此类作用形式多样,包括有效载荷杀伤癌细胞的间接效应(即免疫原性细胞死亡[ICD])、有效载荷对免疫细胞的直接效应、专为激活免疫而设计的有效载荷,以及抗体与免疫细胞的相互作用。上述机制共同构成了ADC作用的基本框架,深入理解这些过程,将有助于推动ADC及其联合疗法的合理设计,从而实现更有效的癌症治疗。
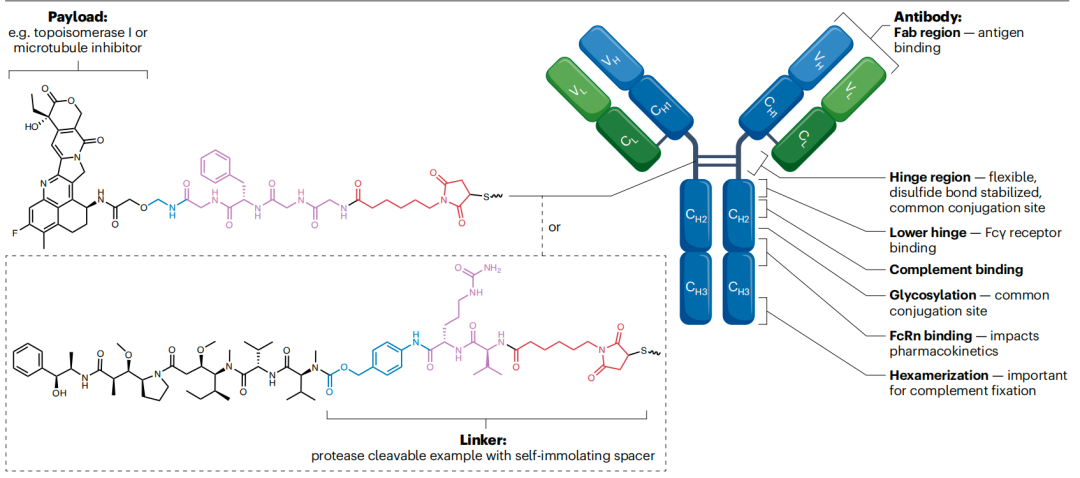
ADC结构
一、ADC的分布与疗效
与所有治疗药物类似,ADC必须从给药部位运输至作用部位。目前获批的ADC均通过静脉注射给药,且必须将其小分子有效载荷递送至TME内的特定细胞位置,例如癌细胞或免疫细胞。这一旅程需克服多重障碍:从体循环、组织渗透到细胞摄取及亚细胞定位,每一阶段都存在独特挑战。
1.系统药代动力学
作为系统性给药疗法,ADC尤其适用于广泛转移性疾病,甚至在脑转移患者中也显示出疗效。经静脉给药后,ADC经血浆迅速分布至全身各组织。由于抗体(约150 kDa)与有效载荷(通常约0.5 kDa)之间的尺寸差异,ADC的分布模式通常与单克隆抗体相似。但早期研究发现,调整抗体上的有效载荷数量会显著改变其分布和清除特性。在抗体表面偶联多个(通常具有亲脂性)有效载荷可能增加与细胞的非特异性相互作用,从而加速系统清除;而有效载荷数量过少又可能导致递送剂量不足,影响细胞杀伤效果。因此,历史上多数ADC的药物抗体比率(DAR)设定为4,旨在平衡疗效与药代动力学特性。
早期的ADC多采用非位点特异性偶联方式,例如将有效载荷连接至赖氨酸上的游离氨基,或通过铰链区二硫键的部分还原实现偶联。当前多数ADC采用铰链区二硫键偶联技术,实现了对铰链区的半位点特异性偶联,避免干扰抗体的功能域。近年来,位点特异性标记和连接子化学的进展改进了ADC偶联技术。这些创新使得可以在抗体上的几乎任何位点进行偶联,尽管仅少数位点能同时满足高稳定性与对药代动力学干扰最小的要求。采用亲水性连接子有助于增强连接子稳定性、改善ADC循环,从而通过精细调节DAR来优化抗体和有效载荷剂量,提升疗效。
有效载荷从ADC释放后,会像小分子药物一样,经历诸如肝脏代谢和排泄等分布与清除过程。体循环中的游离有效载荷主要有两个来源:一是ADC在循环中的解偶联(常见于连接子稳定性较差的ADC);二是ADC被组织摄取后,在细胞内释放有效载荷,继而经内渗作用回流至血液(更常见于连接子稳定性较高的ADC)。目前,体循环中的有效载荷水平与ADC整体抗肿瘤疗效的相关性尚存争议,其影响程度可能取决于每个ADC的具体特征(如连接子稳定性与有效载荷活性)及关键肿瘤相关因素(如靶点表达水平、肿瘤对有效载荷的敏感性)。
随着化学技术的进步,稳定且良性的偶联技术得以实现。ADC的体内分布方式与抗体相似,包括较长的循环时间,并在多种组织中缓慢进行弥散性代谢。由于其较长的循环半衰期,ADC可通过非特异性机制被细胞摄取(如巨胞饮作用),随后被转运至溶酶体释放有效载荷。人体多数细胞表达的新生儿Fc受体(FcRn)能结合ADC的Fc段,并将其循环回细胞表面,从而减少这种非特异性摄取。多数ADC保留了与FcRn的结合能力,使得很多非特异性内吞的ADC能够在表达FcRn的细胞(如内皮细胞这类参与抗体代谢的关键细胞)中重新回到细胞表面。同时,Fc段与其他受体的结合,例如高亲和力Fcγ受体1(FcγRI;亦称CD64)或低亲和力FcγRIIA(亦称CD32A)、 FcγRIIB(亦称CD32B)、FcγRIIIA及FcγRIIIB(亦称CD16A和CD16B),均会影响ADC的系统清除,将其导向不同的免疫细胞群。这种效应在具有不同免疫细胞水平和内源性抗体的动物模型中可能更为显著。Fc结合与抗原特异性Fab结合的共同作用可影响整体清除率,Fc受体聚集会加速这一过程。
ADC的循环半衰期直接影响最终抵达肿瘤的药量。ADC向肿瘤递送的速度限制性步骤是穿过肿瘤血管的外渗过程,因此血管渗透性、肿瘤血管化程度以及ADC的循环时间都会影响最终输送至肿瘤的ADC总量。尽管多数ADC会在全身进行弥散性代谢,但其对癌症抗原的高亲和力有助于将药物保留在肿瘤内部。这种滞留效应将有效载荷富集于肿瘤组织,相较于小分子化疗药物具有显著优势。一旦进入肿瘤,ADC在瘤内的分布情况也对治疗效果具有重要影响。
2.ADC在肿瘤内的分布
ADC在实体瘤内的分布,是影响该类药物治疗效果的一个关键因素。研究表明,抗体在肿瘤中呈异质性分布,而近期基于荧光抗体技术进展,使我们能以细胞级分辨率可视化这种分布差异。在某些肿瘤类型中,例如胰腺癌,物理屏障可能阻碍药物向组织内部渗透。更为普遍的限制来自药代动力学屏障,由于抗体和ADC与靶点的结合速度远快于其向肿瘤内扩散的速度,导致药物更容易滞留在血管周围。增加抗体剂量可使这些近血管区域达到饱和,从而推动药物向肿瘤深层组织渗透。鉴于许多抗体即使在高剂量下仍具有良好耐受性,因此该策略可克服生物制剂的扩散限制。但对ADC而言,有效载荷毒性是决定剂量上限的主要因素。因此,高DAR或超强效的有效载荷会限制ADC的使用剂量,降低其向肿瘤组织的渗透能力,从而削弱治疗效果。
确保抗体剂量充足(与偶联的有效载荷数量或效力无关),是ADC设计的一个重要考量。这也在一定程度上解释了为何获批用于实体瘤的ADC剂量通常高于血液肿瘤——后者面临的药物递送障碍相对较少。
然而,上述增加ADC组织分布的策略存在重要的局限性,过度优化可能适得其反。使用低效力有效载荷(如蒽环类多柔比星)、在临床可耐受的情况下降低DAR以实现肿瘤饱和剂量,或给予过量的未偶联抗体,都可能降低疗效。当肿瘤对ADC的摄取达到饱和后,继续增加剂量并不能进一步提升瘤内递送,这凸显了精细校准DAR以实现最佳临床结果的重要性。
要实现最佳治疗效果,需要采取平衡策略,既要递送足量抗体以覆盖尽可能多的癌细胞,又要保持有效载荷具有足够效力,从而在安全的前提下拓展治疗窗口。
3.ADC在细胞内的处理过程
ADC在肿瘤组织中分布后,需通过细胞摄取与处理才能释放有效载荷。该过程通常始于ADC与癌细胞表面受体结合,随后经内吞作用进入内体-溶酶体通路。在溶酶体中,有效载荷被释放,并通过转运或被动扩散至细胞质或细胞核,进而作用于微管、DNA或拓扑异构酶等特定靶点。ADC的整体疗效受到该通路中多个相互关联且连续步骤的影响。
抗体和ADC能快速结合靶抗原,得益于肿瘤细胞表面抗原的高密度(每细胞1万至≥100万个受体)和结合的高速度。这种结合基本不可逆,因为抗体的高亲和力和/或多价性能够阻止所有Fab段在内吞前与受体解离。
结合后,内吞过程取决于特异性抗体及靶受体的生物学特性。虽然某些具有旁观者效应的有效载荷可通过被动扩散跨膜释放,从而在TME中发挥作用,但对大多数ADC而言,内吞仍是细胞内有效载荷释放的主要途径,通常也更利于高效杀伤细胞。当ADC-受体复合物被内吞后,其在细胞内的运输路径将受抗体特性与受体生物学特性共同影响。随着内体pH值降低,抗体可能与受体解离或保持结合状态。受体随后可能循环回细胞表面,也可能被送至溶酶体降解。解离的抗体可通过与FcRn结合实现再循环,亦可能随其他可溶性蛋白一起被运输至溶酶体。当细胞持续暴露于抗体或ADC时,内吞速率会因体循环与下调而发生动态变化,因此在实验设计中需注意模拟这种持续暴露的环境,以准确反映体内真实过程。
在内体-溶酶体通路中,有效载荷的释放受特定连接子的化学性质调控。最常见类型包括:可水解型(例如戈沙妥珠单抗)、蛋白酶可裂解型(例如维恩妥尤单抗、T-DXd)、二硫键连接型 (例如索米妥昔单抗)、不可裂解型(例如恩美曲妥珠单抗)和葡萄糖醛酸连接子(例如连接曲妥珠单抗与MMAF的ADC)。裂解动力学至关重要:快速裂解有助于药物扩散出细胞(即洗脱现象)前在肿瘤内达到更高的有效载荷浓度。但这必须与连接子在循环中的稳定性相平衡,防止ADC在抵达肿瘤前因蛋白酶、酯酶及水解作用过早释放。
4.旁观者效应与有效载荷分布
有效载荷的细胞膜渗透性,是影响ADC疗效的另一关键因素。尽管大部分有效载荷会在靶细胞内扩散并结合至预定靶点(例如拓扑异构酶I或微管),但具有细胞膜渗透性的有效载荷能够扩散出靶细胞(洗脱),并进入邻近的“旁观者”细胞。这种旁观者效应对于靶点表达异质性的肿瘤非常重要,它使ADC能更有效地作用于肿瘤中抗原低表达和抗原阴性的癌细胞。
虽然旁观者效应可能增加脱靶风险,但不具备此效应的有效载荷同样会导致毒性。当ADC被健康细胞非特异性内吞后,有效载荷因无法洗脱而滞留,从而引发显著毒性反应。与高血浆浓度给药的小分子药物不同,ADC释放的有效载荷总量有限,因此其在亚细胞水平的分布变得至关重要。诸如在细胞膜中的分配、电荷或pKa介导的细胞器分配,以及蛋白质结合等因素,均会显著影响游离有效载荷的渗透距离。即使在有旁观者效应的情况下,旁观者细胞内的药物浓度也远低于直接被ADC靶向的细胞,因此组织渗透性依然是决定疗效的核心。
具有旁观者效应的有效载荷可被动扩散至邻近细胞(包括免疫细胞),而Fc受体的参与能进一步增强这一过程。高亲和力的FcγRI可结合单价抗体,而免疫突触中的受体聚集可驱动与低亲和力FcγRII和FcγRIII的结合。Fc介导的内吞及随后有效载荷释放,可引发显著旁观者效应,即使ADC未直接结合靶向抗原,也能为ADC的疗效做出贡献。FcγRI的结合与内吞效率尤其高,这可能与其在抗体介导的抗原递送至树突状细胞(DC)及其他抗原呈递细胞(APC)中扮演的关键角色有关。将有效载荷递送至免疫细胞或肿瘤基质细胞,可间接杀伤癌细胞。然而临床证据提示,癌细胞直接摄取ADC仍是获得强效治疗应答的主要途径。这可能是因为旁观者杀伤的效率相对较低。
除递送有效载荷外,Fc受体结合还会驱动介导关键免疫功能的信号通路,例如细胞因子释放、抗体依赖性细胞毒性(ADCC)和抗体依赖性细胞吞噬作用(ADCP)。当前ADC采用IgG1亚型以增强效应功能,或选用IgG4亚型以减弱效应功能,两种亚型均具有较长的固有循环半衰期。这些Fc机制已被公认为单克隆抗体疗法起效的重要因素。尽管这些效应能显著增强癌细胞杀伤,但也可能导致脱靶毒性,包括细胞因子释放综合征及高血浆浓度引起的输注相关反应。Fc受体相互作用的多样性与多重效应,很可能是影响疗效与毒性多重表现的原因,这也使其成为设计与应用中必须考量的关键因素。
总之,ADC的设计必须确保在临床剂量下实现充分的循环和肿瘤摄取。其效力与DAR必须平衡,以确保在达到或低于细胞饱和的条件下仍能实现足够杀伤,同时避免因载荷过强而导致毒性过高或肿瘤渗透受限。最后,细胞处理过程必须足以驱动预期的药效学应答,这需要考虑特定肿瘤类型的靶点表达密度和净内吞率。上述每个环节都必须针对具体靶点和ADC技术进行定制,以实现最大的有效载荷递送,并确保有效的组织分布与细胞释放。
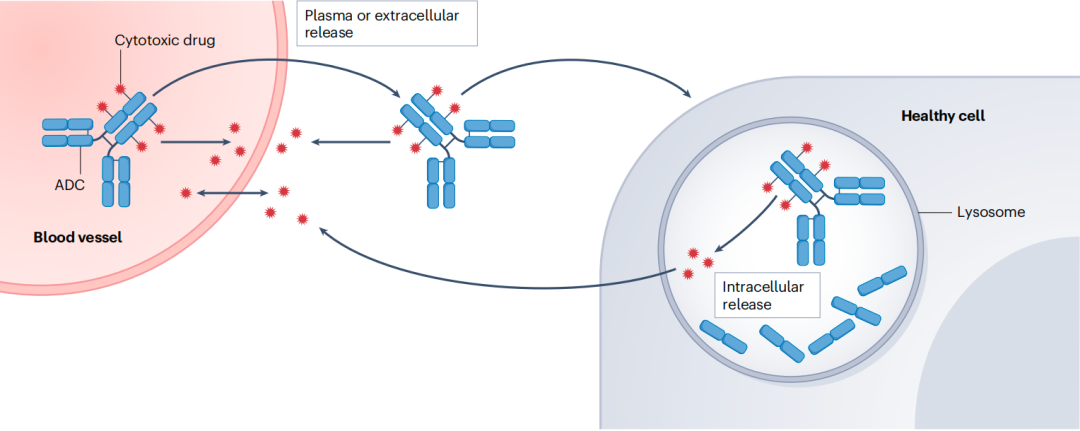
连接子稳定性影响有效载荷向血浆或组织细胞外间隙的释放
有效载荷在血浆或组织细胞外间隙释放(短箭头)
完整ADC外渗、内吞、从细胞内洗脱(弯曲箭头)
游离有效载荷与血浆交换(双箭头)
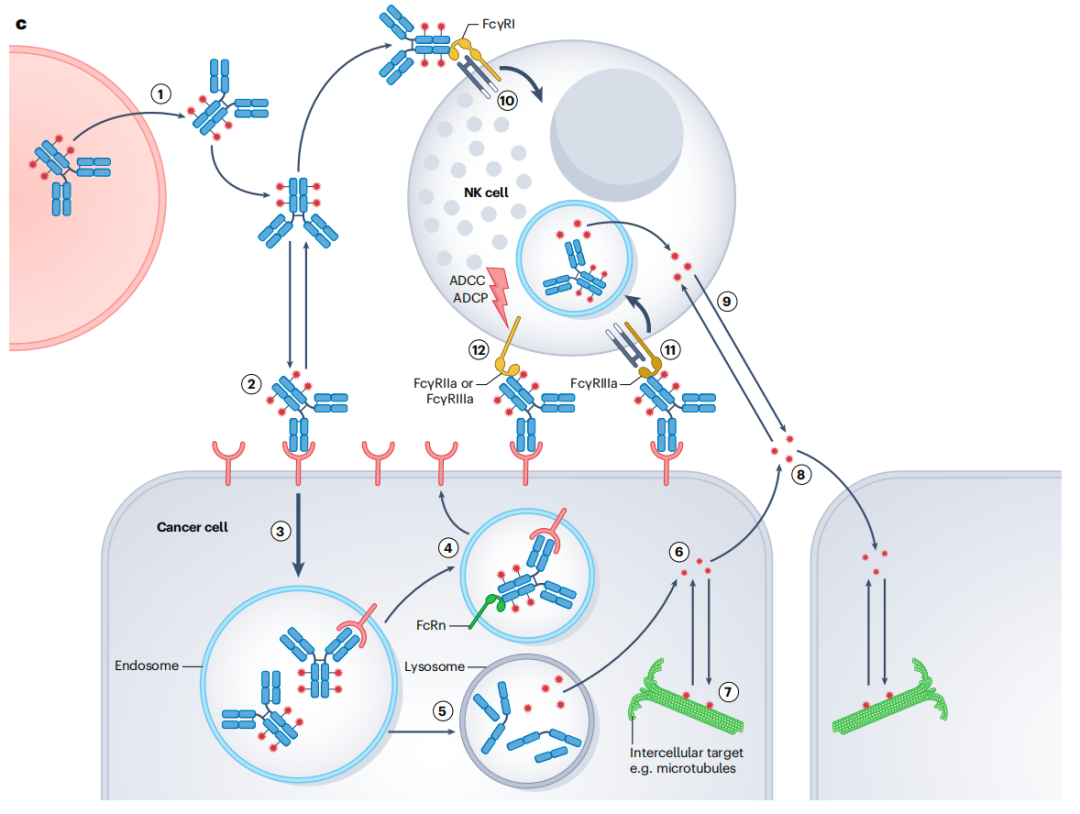
ADC及其有效载荷会与多种细胞相互作用
1.ADC外渗 2.与癌细胞结合 3.内吞 4.再循环回细胞表面 5.导向溶酶体 6.有效载荷溶酶体逃逸 7.与癌细胞内靶点和蛋白质结合 8.旁观者效应 9.有效载荷被动扩散进入免疫细胞 10.ADC结合FcγRI 11.抗体结合使免疫突触中的Fc受体聚集 12.ADCC/ADCP
二、ADC毒性
1.在靶毒性 vs 脱靶毒性
据估计,绝大多数ADC(可能超过总剂量的99%)是在健康细胞内被分解代谢的。尽管如此,与小分子化疗相比,ADC在递送和维持肿瘤内有效载荷浓度方面依然更为高效。这种摄取与代谢过程可能是构成ADC相关毒性的关键驱动因素,或由两种机制介导:一是ADC与靶点受体结合所引发的在靶毒性;二是通过非特异性受体介导的内吞等机制导致的脱靶毒性。此外,有效载荷的旁观者效应在增强疗效的同时,也可能增加毒性风险。
在靶毒性的性质和严重程度,很大程度上取决于靶点受体在健康细胞上的表达水平和内吞动力学。为最大限度降低在靶毒性风险,ADC的研发工作通常侧重于那些在易受化疗毒性影响的健康细胞(如造血干细胞等快速分裂的细胞)上很少或没有表达的细胞表面蛋白。然而,一些ADC毒性仍可能由在靶机制驱动。例如,心脏毒性是接受曲妥珠单抗及基于曲妥珠单抗ADC治疗患者中罕见但严重的副作用,这表明抗体或ADC靶点与HER2阳性心肌细胞的相互作用是此种毒性的主要驱动因素。另一例证是味觉障碍,在维恩妥尤单抗的临床试验中,约40%的患者报告了此副作用。该副作用可能与其靶点(nectin-4)在唾液腺中的表达有关。同样,严重皮疹是Glembatumumab Vedotin的剂量限制性毒性(DLT),可能与该ADC靶点(一种I型跨膜糖蛋白GPNMB,也称为骨激活素)在皮肤中的表达有关。这些特定毒性在携带相同有效载荷但靶向不同抗原的ADC中并不常见,进一步支持了在靶机制。连接子更稳定、有效载荷效力更高的ADC可能会加剧在靶毒性,这凸显了在ADC设计和开发过程中,审慎评估健康组织中靶点表达水平的重要性。
一种克服在靶、非肿瘤毒性的策略是开发双特异性ADC。其设计通常采用“与”门逻辑:对每个靶点的亲和力较低,可减少在单一靶点健康组织上的结合;但当两个靶点在肿瘤细胞上共表达时,可产生亲和力叠加,从而驱动强效摄取并提高对肿瘤的选择性。这不同于对任一靶点均具有高亲和力的“或”门策略。后者旨在提高肿瘤的总靶点密度,扩大ADC潜在受益人群,降低单一靶点下调导致耐药性的风险。随着双特异性蛋白设计的日益复杂,深入理解作用机制对优化药物至关重要。
尽管存在上述案例,但迄今开发的ADC药物中,由靶点介导的健康细胞摄取所引发的DLT仍属罕见。对70项已发表ADC临床研究数据的回顾表明,递送同类有效载荷的ADC均表现出高度相似的毒性谱、DLT和最大耐受剂量,而与其靶向的抗原无关。例如,递送MMAE的ADC均呈现高度相似的DLT,包括败血症、运动神经病变和骨髓毒性;DM1 ADC通常表现出3/4级血小板减少症和肝毒性;而MMAF和DM4 ADC则始终与严重的眼部毒性相关;PBD ADC也呈现高度相似的毒性谱,包括肝酶升高、骨髓抑制及血管渗漏综合征。表明ADC毒性主要由脱靶的有效载荷递送驱动。这种脱靶毒性最终决定了ADC的耐受性和患者用药剂量。
2.ADC有效载荷的脱靶递送:内吞途径
ADC的脱靶毒性既可能源于游离有效载荷的扩散,也可能来自完整ADC被非靶细胞摄取。后者可能通过液相内吞或受体介导的内吞作用发生。单克隆抗体和ADC的非特异性内吞受理化性质影响,高疏水性和正电荷会增加内吞作用。鉴于大多数ADC有效载荷是非极性的,其整体的疏水性随载药量增加而增加。导致高DAR值的ADC通常表现出高系统清除率、低耐受性和窄安全边际。同时,带正电荷的分子易与带负电的细胞膜相互吸引,促进电荷介导的内吞作用。因此,调节ADC的净表面电荷可影响其在正常组织中的非特异性摄取,进而影响毒性。
ADC的Fc段与FcγR结合,引发FcγR介导的内吞及后续毒性。巨噬细胞因高表达FcγR,尤其易通过此途径发生脱靶毒性。例如,间质性肺病(ILD)是多种抗HER2 ADC共有的严重不良事件。虽然具体机制尚未完全明确,但研究显示,T-DXd在灵长类肺泡巨噬细胞中显著富集,而该细胞群所在区域正是ILD病变发生处。鉴于肺泡巨噬细胞高表达FcγR,且呼吸性肺泡上皮HER2表达很低,因此FcγR介导的摄取很可能是诱发ILD的原因。这一推测与多种靶向不同抗原的ADC均可引发ILD的观察结果相符。类似地,巨核细胞通过FcγR介导的内吞作用(特别是FcγRIIa介导),被认为是恩美曲妥珠单抗导致血小板减少症的潜在机制。研究表明,阻断FcγRIIa结合可抑制巨核细胞对恩美曲妥珠单抗的摄取,但也存在相互矛盾的报道。
为深入理解FcγR在ADC毒性中的作用,目前正在临床中评估经改造而降低或消除FcγR结合能力的ADC。若此类药物能有效降低严重副作用(如ILD和血小板减少症)的发生率,则可通过Fc段改造、改变糖基化、可裂解阻断基团及pH敏感性结合位点等多种策略缓解该问题。
此外,甘露糖受体(MR)介导的内吞也被认为是导致ADC脱靶肝毒性的潜在机制。已知MR介导的摄取参与多种治疗性蛋白的清除过程,且随着抗体甘露糖基化水平升高,全身清除率往往同步加快。具体而言,肝窦内皮细胞通过MR摄取ADC,可能是导致ADC非靶点依赖性肝毒性的关键因素。
3.ADC有效载荷的脱靶递送:连接子稳定性与旁观者效应
ADC连接子主要分为可裂解型与不可裂解型。目前超过80%已获批的ADC采用可裂解连接子结合亲脂性有效载荷,通过强效旁观者效应治疗具有异质性抗原表达或抗原内吞速率缓慢的癌症。可裂解连接子的稳定性与ADC毒性之间的关系较为复杂。从药代动力学基本原理看,血浆中释放的有效载荷的累积暴露量取决于其剂量与清除率,理论上与释放速率无关。因此,若毒性直接由释放的有效载荷暴露量决定,那么连接子稳定性差预计并不会显著改变剂量与毒性之间的关系。这或许解释了为何将抗体与有效载荷偶联不会显著改变有效载荷本身的耐受性。然而,连接子稳定性差,预计会降低有效载荷在靶点部位的暴露量(相对于血浆中的暴露量),从而降低靶点部位与非靶点部位细胞毒性的比例,从而缩小ADC的治疗窗口。鉴于系统性有效载荷暴露量可能在ADC毒性中发挥的作用,人们越来越关注使用能结合有效载荷的抗体片段作为ADC的佐剂,以中和血浆中释放的有效载荷,从而减少脱靶毒性并增强ADC的治疗选择性。不过,这类策略仍需临床验证。
与此同时,位点特异性ADC技术在过去几年发展迅速,它通过更精准地调控理化特性,制备出分布更均匀的产品,避免DAR分布不均的问题,旨在通过提升稳定性与优化理化特性来扩大治疗窗口。最新临床证据显示,在若干案例中,位点特异性偶联与非位点特异性偶联的毒性特征存在差异。但需注意,该技术也可能因稳定性增加、暴露延长而加剧某些毒性(如眼部毒性),甚至导致一些在研ADC终止开发。因此,位点特异性偶联技术虽提供了更精细的调控手段,但要实现更优的治疗窗口,仍需深入理解其多维度的毒性特征。
另一种降低毒性的思路是开发耐受性更好的有效载荷(例如蛋白质降解剂、抗凋亡蛋白BCL-xL抑制剂、酪氨酸激酶抑制剂等)。但主要挑战在于,这些载荷需具有足够效力以介导药效学效应。由于ADC的特异性摄取受限于受体表达和内吞过程,其最大递送能力有限。但扩展合适的有效载荷对ADC领域仍至关重要。除降低毒性外,对避免序贯使用同类载荷产生交叉耐药也具有重要意义。
总体而言,绝大多数给药的ADC并未到达肿瘤,而是被健康组织通过多种途径摄取。释放的有效载荷在局部组织的浓度以及特定组织对有效载荷的敏感性,很大程度上决定了ADC的毒性特征和最大耐受剂量。鉴于有效载荷是毒性的主要决定因素,且通常与抗体无关,因此在实现治疗效果与维持临床可耐受的总有效载荷剂量之间取得平衡至关重要。
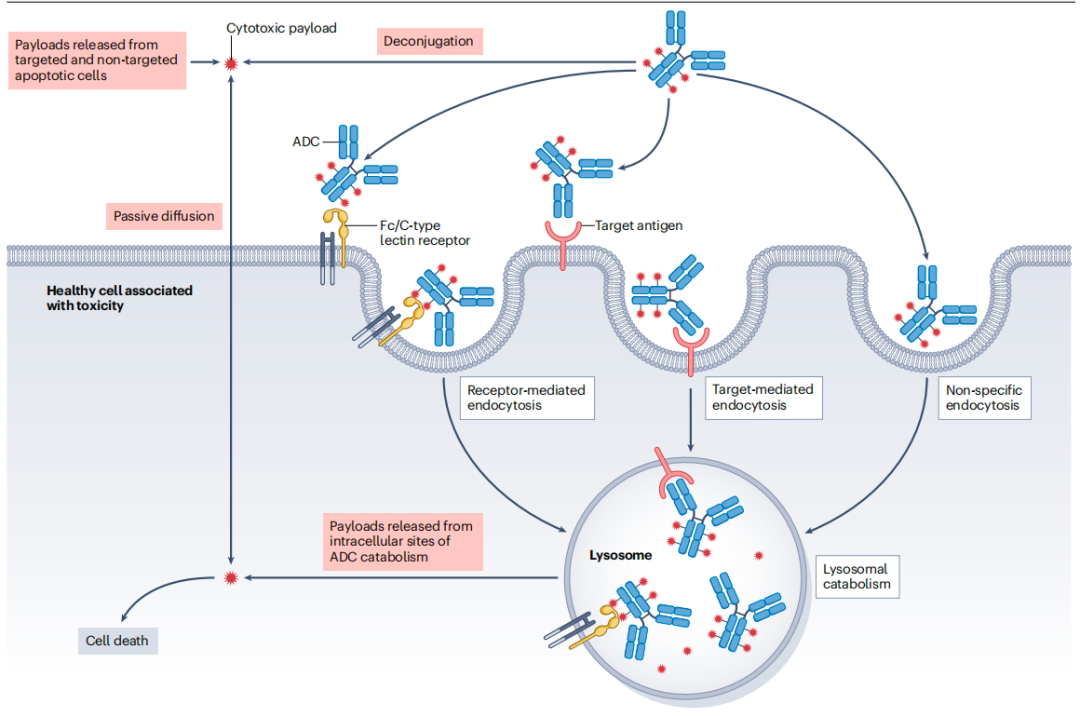
导致ADC毒性的机制
三、ADC的免疫调节作用
尽管ADC主要通过有效载荷诱导癌细胞死亡,但其独特设计还能引发强烈的免疫应答,从而将免疫肿瘤学与ADC研发紧密联系起来。因此,了解ADC治疗如何影响抗肿瘤免疫应答,对优化其治疗潜力至关重要。
免疫系统通过持续监测并清除恶性细胞来防止癌症发生。其中,细胞毒性CD8⁺ T细胞能特异性识别肿瘤抗原,并建立持久的免疫记忆,这是适应性免疫系统的标志性优势。然而,强效且持久的抗肿瘤免疫力同样依赖于先天免疫系统,包含中性粒细胞、自然杀伤(NK)细胞以及APC,如巨噬细胞和DC。
先天免疫细胞可被胚系编码的模式识别受体(PRR)激活,这些受体能识别垂死肿瘤细胞释放的损伤相关分子模式(DAMP)。DAMP能促进APC的聚集与成熟,使其能够吞噬肿瘤相关抗原并呈递给初始T细胞,从而启动适应性抗肿瘤T细胞应答。此外,DAMP还能促使DC和巨噬细胞等先天免疫细胞释放促炎性细胞因子(尤其是I型干扰素),进而增强TME内的抗原呈递、免疫细胞聚集,以及NK细胞和CD8⁺ T细胞的活化。
DC以其卓越的抗原呈递和T细胞激活能力,在抗肿瘤免疫的启动与调控中处于核心地位。然而,在TME中,DC的成熟过程常常受到抑制,导致其活性减弱,进而引发T细胞应答不足。因此,克服这些抑制机制,成为增强抗肿瘤T细胞免疫的重要方向。
1. 细胞毒性有效载荷与ICD
携带细胞毒性有效载荷的ADC展现出显著的免疫调节潜力。在免疫功能不全的临床前模型中,其疗效往往下降,提示免疫调节很可能也是其主要的作用机制之一。虽然免疫调节对ADC疗效的具体贡献仍在探索中,但持续的研究正不断揭示ADC与免疫通路相互作用的新机制。
ICD在免疫监视中发挥关键作用。与其他能逃避免疫检测的细胞死亡形式不同,ICD能促使垂死的癌细胞有序释放DAMP,如钙网蛋白(CRT)、ATP和高迁移率族蛋白B1(HMGB1),从而启动抗肿瘤免疫应答。在ICD早期,CRT会转运至细胞膜并作为“吞噬我”信号,促进DC及其他APC的吞噬作用。随着细胞死亡进程推进,ATP作为趋化因子,引导APC等免疫细胞向死亡部位聚集,并促进促炎性细胞因子的释放。最终,HMGB1进一步增强免疫细胞的聚集,并持续刺激促炎反应。这种由DAMP驱动的过程可激活肿瘤特异性T细胞,从而在抗原特异性抗肿瘤免疫中发挥核心作用。
在临床前模型中,多种携带细胞毒性有效载荷的ADC,均可诱导ICD,包括CRT、ATP和HMGB1的释放。这些研究揭示了ICD诱导的DAMP释放与广泛免疫激活及肿瘤控制之间的关联。例如,蒽环类ADC T-PNU在HER2阳性乳腺癌小鼠模型的TME中诱导了显著的免疫基因组学改变,并增强了CD8⁺ T细胞的活化。重要的是,这类药物触发的免疫应答能在后续的肿瘤再攻击中抑制肿瘤生长,表明ICD可促进长期的免疫监视。因此,能诱导ICD的ADC具备启动肿瘤特异性T细胞应答的能力,为其与ICI的联合应用提供了理论依据。事实上,临床前研究已证实,能诱导ICD的ADC与ICI可产生协同效应,印证了二者在机制上的互补性。
2. 细胞毒性有效载荷对DC的直接激活
部分细胞毒性有效载荷可以不依赖ICD,直接激活DC。尽管DC通常不会内吞完整的ADC,但能对TME中游离的有效载荷做出反应。研究表明,靶向微管的药物能以不同效力广泛诱导DC成熟与活化,从而增强其在肿瘤部位的抗原摄取能力,并促进其向肿瘤引流淋巴结迁移。虽然微管解聚剂和微管稳定剂在临床相关剂量下,主要通过相似机制抑制微管动力学、诱导有丝分裂停滞,但在临床前模型中,免疫刺激效应主要见于微管解聚剂。机理研究发现,微管失稳所引发的DC成熟依赖于GEF-H1,该蛋白RHO通路的关键激活因子。这些发现为进一步研究抗肿瘤免疫中微管网络与信号传导通路之间复杂的相互作用奠定了基础。
直接激活DC的能力并不限于微管解聚剂类有效载荷。在同源小鼠模型中,含拓扑异构酶I抑制剂的ADC同样能诱导DC成熟,增强抗肿瘤免疫,并对肿瘤的再次攻击产生保护作用。与能诱导ICD的ADC类似,大量研究证实,能激活DC的ADC与ICI联合使用时可发挥协同效应。
尽管在体外试验中,微管解聚剂相较于拓扑异构酶抑制剂,展现出更强的DC激活能力,但在ADC治疗背景下,这些差异与临床的相关性尚不明确。
3. Fc段的免疫调节作用
IgG抗体的Fc段能启动多种效应功能,包括ADCC、ADCP、抗原呈递,以及趋化因子和细胞因子的分泌。在设计Fc效应功能时,选择位于肿瘤细胞膜附近的靶抗原表位至关重要,这种空间上的邻近性有利于受体聚集,并通过物理性排斥CD45等大型磷酸酶,促进更有效的免疫突触形成,从而优化FcγR介导的免疫细胞活化——这与传统ADC设计中优先考虑的高效内吞特性有所不同。诸如利妥昔单抗、曲妥珠单抗和西妥昔单抗等多种治疗性抗体均可诱导ADCC。在介导ADCC的过程中,自然杀伤(NK)细胞因其固有的细胞毒性和高表达活化型FcγR而尤为关键。临床前模型表明,ADC仍能保留这些抗体介导的效应功能。例如,曲妥珠单抗与DM1或DXd偶联后,其在乳腺癌细胞中诱导ADCC的能力并未减弱。此外,通过改造Fc段可增强或削弱其与FcγR的结合能力,进而调控其免疫调节潜力。
ADC的Fc段功能不仅限于激活经典的免疫效应应答。研究意外地发现,通常被认为促癌的肿瘤相关巨噬细胞(TAM)也可能通过FcγR介导的途径,以非靶点依赖的方式摄取ADC,进而对周围肿瘤细胞产生“旁观者杀伤”效应,并贡献于ADC的整体疗效。这一机制为在抗原异质性或低表达肿瘤中增强疗效提供了可能。然而,非靶点依赖性的摄取与代谢过程也可能导致对非恶性细胞的杀伤,从而引发脱靶毒性。尽管目前尚不明确肿瘤特异性T细胞是否直接受影响,但其潜在的免疫抑制作用凸显了持续研究的必要性,以在优化ADC疗效的同时减轻毒性。
4.免疫刺激抗体偶联物(ISAC)
与传统ADC不同,ISAC携带的是PRR激动剂,能特异性激活TME中的先天免疫应答,从而建立强劲持久的T细胞免疫。相较于可能引发严重脱靶毒性的全身性PRR激动剂,ISAC有望提供一种更精准的方案。
Toll样受体(TLR)是重要的PRR家族,可识别DAMP启动炎症和防御机制。激活位于APC内体的TLR能促进细胞成熟并诱导促炎细胞因子产生,从而增强抗肿瘤免疫。临床前数据显示,靶向APC内TLR7、TLR8和TLR9的ISAC,能有效激发抗肿瘤免疫应答。尽管部分此类ISAC因毒性等问题在临床开发中受阻,但通过优化靶点与激动剂组合,该策略仍在持续探索中。
cGAS-干扰素基因刺激因子(STING)通路是先天免疫激活的核心组成部分,可响应来自病原体或垂死肿瘤细胞的胞质双链DNA。无论在先天免疫细胞还是肿瘤细胞中,激活胞质内的STING均会诱导I型干扰素和其他促炎细胞因子的产生,从而发挥抗肿瘤效应。然而,尽管具有抗肿瘤潜力,游离的STING激动剂仍面临化合物稳定性差与毒性大等多重局限。利用ISAC实现STING激动剂的靶向递送,有望克服这些障碍。
临床前研究表明,靶向EGFR的STING激动剂ISAC,能在显著低于游离药物的剂量下,于肿瘤细胞和髓系细胞中同时激活STING,显著抑制肿瘤生长。值得注意的是,在肿瘤细胞内特异性激活STING可诱导III型干扰素产生,进而促进I型干扰素及其他促炎细胞因子的分泌,显著增强抗肿瘤免疫应答。这些发现揭示了肿瘤自身STING激活与III型干扰素在STING介导的抗肿瘤应答中,可能存在被低估的作用机制。尽管此类ISAC在临床前模型中与其他疗法联用展现出协同效应及完全肿瘤消退的潜力,但早期临床试验中也出现了严重毒性事件,凸显了精确调控给药以平衡疗效与安全性的至关重要性。
5.ADC联合疗法的合理设计
过去十年间,ADC与ICI的获批数量均快速增长。临床前和临床证据表明,合理联合其他抗癌药物,可进一步增强ICI的疗效,实现抗肿瘤活性的叠加或协同效应。化疗是目前较成熟的联合策略,其不仅可诱导免疫原性细胞死亡,还能调节免疫微环境,增强抗肿瘤免疫。相较传统化疗,ADC凭借其靶向递送特性,有望成为ICI更具特异性且更有效的联合伙伴。
然而,ADC的免疫刺激作用在临床中尚未完全明确。不同研究对其能否增强T细胞活化、提高免疫细胞浸润等问题的结论并不一致,部分联合疗法的临床试验结果亦存在矛盾。这提示我们,需进一步探索ADC在真实临床环境中的免疫调节潜力。
目前仅少数ADC-ICI联合方案获批或进入临床后期阶段。尽管早期数据显示部分联合策略具有抗肿瘤
引用格式:ADC在癌症治疗中的分子与免疫学驱动机制. 发布日期:1766655066. 来源:TalkMED拓麦. URL: https://portal.talkmed.com/news/3196
分享
收藏
点赞


